INDICE ARTICOLO
Turnover delle Fibre Elastiche
Le fibre elastiche, come tutte le proteine della matrice extracellulare, hanno una notevole longevità e la loro vita è misurata in anni.
Questa caratteristica predispone le fibre elastiche a un rischio maggiore di invecchiamento molecolare per l’accumulo di danni ripetuti, mentre le proteine intracellulari vivono per ore o massimo giorni.
Per esempio, si stima che il collagene I e II abbia una vita media tra i 15 e i 95anni, mentre l’emivita delle fibre elastiche può arrivare all’età dell’individuo.
La sintesi e il deposito di elastina è prevalentemente limitato al periodo fetale e post-natale, invece negli adulti si verifica un decadimento dell’mRNA dell’elastina.
Danni alle Fibre Elastiche
Per quanto detto, ogni proteina delle fibre elastiche è tenuta a funzionare per molti anni e a resistere all’accumulo di danni da legami crociati mediati dal glucosio, all’accumulo di lipidi e calcio, e alle modifiche tempo-dipendenti nei residui di acido aspartico.
Questi eventi hanno un profondo effetto nel rimodellamento dei tessuti ricchi di elastina.
Gli studi recenti sull’invecchiamento cellulare sono concentrati sulla regolazione e attivazione delle MMP ovvero le metalloproteasi (o metalloproteinasi) che hanno il compito di degradare la matrice extracellulare.
Effetto dei Raggi Solari
L’azione delle MMPs è incrementata dall’esposizione ai raggi UV ed è in grado di catabolizzare componenti delle fibre elastiche come fibrillina-1 e fibulina-5. Questi enzimi hanno però una bassa substrato-specificità che li rende meno efficaci.
Studi hanno confermato che il collagene I è resistente ai cambiamenti strutturali a dosi di UVR raggiungibili fisiologicamente, mentre le microfibrille ricche di fibrillina e il collante tra glicoproteine e fibronectine sono profondamente alterati dalle stesse dosi di radiazioni ultravioletti.
La differente sensibilità può essere dovuta alla composizione aminoacidica diversa. Infatti, le fibre elastiche associate a proteine, tranne l’elastina, sono ricche di cromofori UVB (Cys, His, Phe, Tyr e Trp) che assorbono queste radiazioni.
Si può quindi affermare che le fibre elastiche sono sensibili alla degradazione diretta degli UV con conseguenze nell’omeostasi del tessuto.
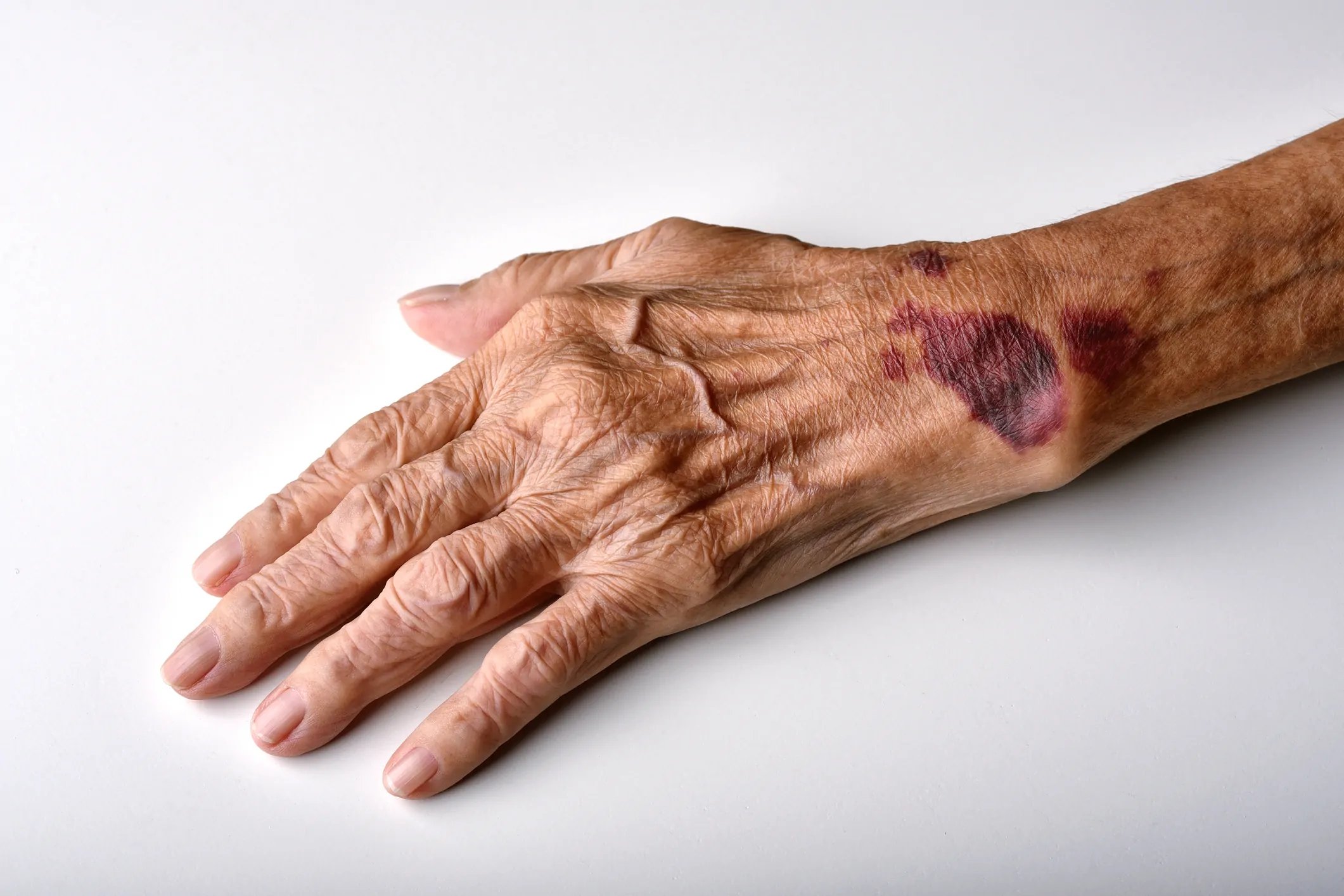
I processi intrinseci ed estrinseci d’invecchiamento causano una perdita di conformità e resistenza del tessuto, e la conseguente formazione di elastosi e rughe.
Conseguenze
Il rimodellamento delle fibre elastiche ha quindi profondi effetti biochimici e cellulari. Frammenti di elastina (detta anche elastochina) possono influenzare il sistema immunitario, agendo sui linfociti per regolare l’espressione dell’elastasi e promuovere l’apoptosi, e sui monociti per contrastare gli stimoli pro-infiammatori.
I peptidi di elastina e fibrillina-1 influenzano il fenotipo di cellule non-infiammatorie inducendo l’espressione di MMPs. Per esempio l’elastina induce l’attivazione delle MMP di membrana di tipo 1 e 2 (MT1-MMP, MMP-2), mentre la fibrillina-1 regola l’espressione delle MMP-1 e MMP-3.
Tuttavia, alla fine, tutte le MMPs hanno la capacità di degradare le maggiori proteine componenti la EMC e i componenti della DEJ.
Da studi recenti si evince che le microfibrille di fibrillina sono importanti anche per l’omeostasi del tessuto perché sequestrano TGFß.
Il TGFß lega intercellularmente famiglie di proteine latenti legate ad esso formando così un complesso proteico che, a sua volta, si associa extracellularmente a microfibrille ricche di fibrillina.
In definitiva il sequestro di TGFß, l’accumulo di danni nel corso del tempo, i meccanismi degradativi cellulari (MMP) e acellulari (UVR) sono capaci di influenzare profondamente l’architettura, quindi il rimodellamento delle fibre elastiche, e avere anche effetti sulle funzioni biochimiche delle molecole.
Conseguenze strutturali e funzionali del rimodellamento della matrice extracellulare dermicaTrattamenti per le modificazioni del sistema di Fibre Elastiche del Derma